Memory Improves Dramatically Between 1 and 2 Months of Age and Then Again by 36 Months
The Development of Attending Systems and Working Memory in Infancy
What are the mechanisms that support the ability to retain information for a period of time before interim on it? When does this ability sally in human evolution? What role does the development of attention play in this procedure? Answers to these questions are non only important for furthering our understanding of working memory, but are also cardinal to understanding cognitive development at a broader level. We delve into these questions from a developmental cerebral neuroscience perspective with a particular focus on the touch on of the development of attention systems on recognition retentiveness and working retention. In the sections that follow, nosotros nowadays a selective review of research in which psychophysiological and neuroscience techniques accept been combined with behavioral tasks to provide insight into the effects of babe attending on performance on recognition memory tasks. Nosotros begin our review with a focus on babe attending and recognition memory considering the combined measures used in this line of piece of work provide unique insight into the influence of sustained attention on memory. To date, this approach has yet to be utilized to examine relations between attention and working memory in early development. In the 2d half of the article, nosotros review enquiry on working retentiveness in infancy with a focus on studies utilizing behavioral and neuroscience measures (for more exhaustive reviews, run across Cowan, 1995; Nelson, 1995; Pelphrey and Reznick, 2003; Rose et al., 2004; Bauer, 2009; Rovee-Collier and Cuevas, 2009). We also focus on contempo enquiry findings that shed light on neural systems potentially involved in attention and working memory in infancy (for excellent reviews on attention and working memory relations in childhood, come across Astle and Scerif, 2011; Amso and Scerif, 2015). Because the human infant is incapable of producing exact or complex behavioral responses and also cannot be given instructions on how to perform a given job, past necessity, many of the existing behavioral studies on infant working memory accept been built upon look elapsing or preferential looking tasks traditionally used to tap into babe visual attention and recognition retentivity. Thus, it is hard to depict singled-out lines when determining the relative contribution of these cerebral processes to performance on these tasks in the infancy menstruation (but see Perone and Spencer, 2013a,b). We conclude with a section examining potential relations between attending and working retentivity and suggest that the development of attention systems plays a primal role in the timing of pregnant gains in working retentivity observed in the second half of the first postnatal year.
Infant Visual Attention and Recognition Memory
Much of what we know about the early development of visual attending comes from a large torso of enquiry on recognition memory in infancy. Because the defining feature of recognition retentivity is differential responsiveness to novel stimuli in comparison to familiar (or previously viewed) stimuli (Rose et al., 2004), the bulk of behavioral enquiry in the area has utilized the visual paired comparing (VPC) task. This chore involves the simultaneous presentation of two visual stimuli. Expect duration to each stimulus during the paired comparing is measured. Nether the framework of Sokolov's (1963) comparator model, longer looking to a novel stimulus in comparison to a familiar stimulus (i.e., a novelty preference) is indicative of recognition of a fully encoded familiar stimulus. In contrast, familiarity preferences are indicative of incomplete processing and continued encoding of the familiar stimulus. The underlying assumption is that infants will continue to look at a stimulus until it is fully encoded, at which betoken attention will be shifted toward novel information in the surrounding environment.
Thus, infant look elapsing has been a widely used and highly informative behavioral mensurate of baby attention that besides provides insight into retentivity in early on development. Findings from these studies signal that older infants require less familiarization time to demonstrate novelty preferences than younger infants; and inside age groups, increasing the amount of familiarization results in a shift from familiarity preferences to novelty preferences (Rose et al., 1982; Hunter and Ames, 1988; Freeseman et al., 1993). Older infants also evidence evidence of recognition with longer delays between familiarization and testing. For example, Diamond (1990) establish that 4-month-olds demonstrate recognition with upwards to 10 due south delays between familiarization and testing, half-dozen-calendar month-olds demonstrate recognition with up to 1 min delays, and 9-month-olds demonstrate recognition with up to x min delays. These findings bespeak that with increasing age, infants are able to process visual stimuli more efficiently and afterwards recognize those stimuli after longer delays. Unfortunately for infancy researchers, look duration and attending are not isomorphic. For example, it is not uncommon for infants to go on looking at a stimulus when they are no longer actively paying attention; therefore, looking measures lone practise not provide a particularly authentic measure out of infant attention. This phenomenon is most prevalent in early on infancy and has been referred to as attention capture, obligatory attention, and sticky-fixation (Hood, 1995; Ruff and Rothbart, 1996).
Richards and colleagues (Richards, 1985, 1997; Richards and Casey, 1992; Courage et al., 2006; for review, Reynolds and Richards, 2008) take utilized the electrocardiogram to place changes in eye charge per unit that coincide with different phases of infant attention. During the class of a single expect, infants volition bike through iv phases of attending—stimulus orienting, sustained attention, pre-attending termination, and attention termination. The most relevant of these phases are sustained attention and attending termination. Sustained attention is manifested as a meaning and sustained decrease in eye rate from prestimulus levels that occurs when infants are actively engaged in an attentive state. Attending termination follows sustained attention and is manifested as a return of eye rate to prestimulus levels. Although the infant is notwithstanding looking at the stimulus during attention termination, she/he is no longer engaged in an attentive land. Infants crave significantly less time to process a visual stimulus if heart charge per unit is measured online and initial exposure is given during sustained attending (Richards, 1997; Frick and Richards, 2001). In stark contrast, infants given initial exposure to a stimulus during attention termination exercise not demonstrate evidence of recognition of the stimulus in subsequent testing (Richards, 1997).
The General Arousal/Attention System
Richards (2008, 2010) has proposed that sustained attention is a component of a general arousal organization involved in attending. Areas of the brain involved in this general arousal/attention organisation include, the reticular activating system and other brainstem areas, thalamus, and cardio-inhibitory centers in frontal cortex (Reynolds et al., 2013). Cholinergic inputs to cortical areas originating in the basal forebrain are as well involved in this system (Sarter et al., 2001). Activation of this organization triggers cascading furnishings on the overall state of the organism which foster an optimal range of arousal for attending and learning. These effects include: decreased heart rate (i.east., sustained attention), motor quieting, and release of acetylcholine (ACh) via corticopetal projections. Ruff and Rothbart (1996) and Ruff and Capozzoli (2003) description of "focused attention" in children engaged in toy play as being characterized by motor quieting, decreased distractibility, and intense concentration coupled with manipulation/exploration would be considered a behavioral manifestation of this full general arousal/attention system.
The general arousal/attention system is functional in early on infancy but shows considerable development across infancy and early childhood with increased magnitude of the 60 minutes response, increased periods of sustained attention, and decreased distractibility occurring with increasing historic period (Richards and Cronise, 2000; Richards and Turner, 2001; Reynolds and Richards, 2008). These developmental changes almost likely have a direct influence on performance on working memory tasks. The general arousal/attention system is non-specific in that it functions to modulate arousal regardless of the specific job or function the organism is engaged in. The effects of the system on arousal and attention are as well full general and do not vary in a qualitative manner depending on cognitive chore, thus sustained attention would be expected to influence recognition retentivity and working memory in a like manner. This non-specific attention system directly influences functioning of three specific visual attention systems that also show considerable development in the infancy period. These specific attending systems are: the reflexive system, the posterior orienting system, and the anterior attending system (Schiller, 1985; Posner and Peterson, 1990; Johnson et al., 1991; Colombo, 2001).
The Development of Attention Systems in the Brain
At birth, newborn visual fixation is believed to be primarily involuntary, exogenously driven, and exclusively under the control of a reflexive system (Schiller, 1985). This reflexive arrangement includes the superior colliculus, the lateral geniculate nucleus of the thalamus, and the master visual cortex. Many newborn fixations are reflexively driven by directly pathways from the retina to the superior colliculus (Johnson et al., 1991). Baby looking is attracted past bones only salient stimulus features processed via the magnocellular pathway that tin can more often than not be discriminated in the peripheral visual field, such as high-contrast borders, motion, and size.
Looking and visual fixation stays primarily reflexive for the starting time two months until the cease of the newborn menstruation when the posterior orienting organisation reaches functional onset. The posterior orienting system is involved in the voluntary control of center movements, and shows considerable development from 3 to 6 months of age. Areas of the brain involved in the posterior orienting system include: posterior parietal areas, pulvinar, and frontal eye-fields (Posner and Peterson, 1990; Johnson et al., 1991). The posterior parietal areas are believed to be involved in disengaging fixation and the frontal heart-fields are primal for initiating voluntary saccades. In back up of the view that the ability to voluntary disengage and shift fixation shows significant development beyond this historic period range, Effigy 1 shows results from a expect elapsing study by Courage et al. (2006) in which babe look elapsing dropped significantly to a broad range of stimuli from 3 to 6 months of age (i.e., 14–26 weeks of age).
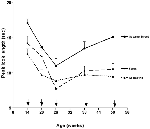
Figure 1. Mean peak look durations for faces, geometric patterns, and Sesame Street as a function of historic period (figure adapted from Courage et al., 2006). Arrows indicate verbal examination age.
At around 6 months of historic period, the anterior attention system reaches functional onset and infants begin the drawn out process of developing inhibitory command and higher guild attentional command (i.east., executive attention). Not only exercise infants have better voluntary control over their visual fixations, they can at present inhibit attention to distractors and maintain attending for more than prolonged periods when it is called for. Equally can exist seen in Figure 1, Courage et al. (2006) found that from vi to 12 months of age (i.e., 20–52 weeks), infants continue to show brief looks to bones, geometric patterns only begin to prove longer looking toward more complex and engaging stimuli such as Sesame Street or human faces. This indicates the emergence of some rudimentary level of attentional control at around 6 months of historic period. Given that several models emphasize some aspect of attentional control equally a core component of working memory (e.g., Baddeley, 1996; Kane and Engle, 2002; Klingberg et al., 2002; Cowan and Morey, 2006; Astle and Scerif, 2011; Amso and Scerif, 2015), it stands to reason that the emergence of attentional control at around 6 months of historic period would contribute significantly to the development of working memory.
The theoretical models for the attending systems discussed above are largely based on findings from comparative research with monkeys, adult neuroimaging studies, or symptomology of clinical patients with lesions to certain areas of the encephalon. Unfortunately, developmental cognitive neuroscientists are highly limited in non-invasive neuroimaging tools available for apply in basic scientific discipline with baby participants. However, nosotros have conducted multiple studies utilizing event-related potentials (ERPs) forth with heart charge per unit measures of attention and behavioral measures of recognition memory (Reynolds and Richards, 2005; Reynolds et al., 2010). Findings from these studies provide insight into potential areas of the brain involved in attention and recognition memory in infancy.
The ERP component which is most conspicuously related to infant visual attention is the Negative central (Nc) component. The Nc is a high amplitude, negatively-polarized component that occurs from 400 to 800 ms postal service stimulus onset at frontal and midline leads (see Effigy 2). Nc has been plant to be greater in amplitude to: oddball compared to standard stimuli (Courchesne et al., 1981), novel compared to familiar stimuli (Reynolds and Richards, 2005), mother'due south face compared to a stranger's face up (de Haan and Nelson, 1997), and a favorite toy compared to a novel toy (de Haan and Nelson, 1999). These findings betoken that regardless of novelty or familiarity, Nc is greater in amplitude to the stimulus that grabs the infant's attention the nigh (Reynolds et al., 2010). Additionally, Nc is greater in amplitude when infants are engaged in sustained attending (as measured by heart rate) than when infants accept reached attending termination (Richards, 2003; Reynolds et al., 2010; Guy et al., in printing). The Nc is likewise ubiquitous in ERP research utilizing visual stimuli with infant participants. Taken together, these findings signal that Nc reflects amount of attentional engagement.
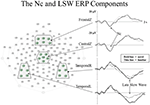
Figure 2. Upshot-related potential (ERP) waveforms and electrode locations for the Nc and late deadening wave (LSW) ERP components. The ERP waveforms are shown to the right. Change in amplitude of the ERP from baseline values is represented on the Y-axis, and fourth dimension following stimulus onset is represented on the Ten-axis. The electrode locations for each of the waveforms are shown to the left in boxes on the layout of the EGI 128-channel sensor net (figure adapted from Reynolds et al., 2011).
In order to determine the cortical sources of the Nc component. Reynolds and Richards (2005) and Reynolds et al. (2010) conducted cortical source analysis on scalp-recorded ERP. Cortical source assay involves computing a forward solution for a prepare of dipoles, and comparison the false topographical plots produced by the forward solution to the topographical plots obtained from observed data. The forrad solution is iterated until the best fitting solution is found. The results of the cortical source analysis can then exist mapped onto structural MRIs. Figure 3 shows the results of our source analysis of the Nc component measured during brief stimulus ERP presentations and also during performance of the VPC task. As tin be seen in Figure 3, the cortical sources of the Nc were localized to areas of prefrontal cortex (PFC) for all age groups including four.v-month-olds. Areas which were common dipole sources included inferior and superior PFC, and the anterior cingulate. The distribution of the dipoles as well became more than localized with increasing historic period. These findings support the proposal that PFC is associated with infant attention, and betoken that at that place is overlap in brain areas involved in both recognition retentivity and working memory tasks. Neuroimaging research with older children and adults indicates that in that location is a neural circuit including parietal areas and PFC involved in working memory (e.g., Goldman-Rakic, 1995; Fuster, 1997; Kane and Engle, 2002; Klingberg et al., 2002; Crone et al., 2006).
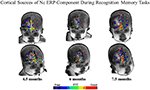
Figure 3. Common equivalent current dipoles activated across recognition memory tasks. Age groups are divided into separate columns. The all-time fitting areas in common between the ERP and visual paired comparing (VPC) tasks are indicated using the color scale. The majority of best fitting areas were located in inferior prefrontal regions (effigy adapted from Reynolds et al., 2010).
The late slow wave (LSW) ERP component is associated with recognition retentivity in infancy. The LSW shows a reduction in amplitude with repeated presentations of a single stimulus (de Haan and Nelson, 1997, 1999; Reynolds and Richards, 2005; Snyder, 2010; Reynolds et al., 2011). As shown in the two lower ERP waveforms in Effigy ii, the LSW occurs from about 1–2 s mail stimulus onset at frontal, temporal, and parietal electrodes. By examining the LSW, Guy et al. (2013) found that individual differences in infant visual attention are associated with utilization of dissimilar processing strategies when encoding a new stimulus. Infants who tend to demonstrate cursory but broadly distributed fixations (referred to every bit short lookers; e.thousand., Colombo and Mitchell, 1990) during exposure to a novel stimulus after showed evidence of discriminating hierarchical patterns based on changes in the overall configuration of private elements (or local features). In contrast, infants who tend to demonstrate longer and more narrowly distributed visual fixations (referred to as long lookers) showed prove of discriminating patterns based on changes in local features merely not based on changes in the overall configuration of local features. Furthermore, research utilizing centre rate measures of attention during functioning on a recognition memory ERP task have provided informative findings regarding relations between attention and memory. Infants are more likely to demonstrate differential responding to familiar and novel stimuli in the LSW when heart rate indicates they are engaged in sustained attention (Richards, 2003; Reynolds and Richards, 2005).
No studies to date have utilized cortical source assay to examine cortical sources of the LSW. Late-latency and long elapsing ERP components can exist more than problematic for cortical source analysis due to greater variability in the timing of the latency of the component across participants and trials, and the likely contribution of multiple cortical sources to the ERP component observed in the scalp-recorded EEG. However, research with non-human primates and neuroimaging studies with older children and adults indicates the function of a medial temporal lobe circuit in recognition retentiveness processes. Cortical areas involved in this circuit include the hippocampus and parahippocampal cortex; entorhinal and perirhinal cortices; and the visual area TE (Bachevalier et al., 1993; Begleiter et al., 1993; Fahy et al., 1993; Li et al., 1993; Zhu et al., 1995; Desimone, 1996; Wiggs and Martin, 1998; Xiang and Brownish, 1998; Wan et al., 1999; Brown and Aggleton, 2001; Eichenbaum et al., 2007; Zeamer et al., 2010; Reynolds, 2015). Regardless of the potential areas involved in recognition memory in infancy, attention is clearly an integral component of successful performance on recognition memory tasks. Performance on recognition memory tasks is influenced by the development of each of the attending systems described in a higher place and it stands to reason that these attention systems would influence performance on working retention tasks in a similar fashion. Furthermore, working memory and recognition memory are closely related and some of the tasks used to mensurate maintenance of items in working memory (i.eastward., visual brusque term memory, VSTM) in infancy are slightly modified recognition retention tasks. Thus, distinctions between working memory and recognition memory tin can be particularly difficult to make during the infancy menses.
The Development of Working Retention in Infancy
Similar to work on attention and recognition memory, research on the early on development of working memory has focused on the use of behavioral measures (looking and reaching tasks) with infant participants. Neuroscience models of early working memory development have as well largely relied on findings from comparative research, clinical cases, and neuroimaging with older children and adults. All the same, in that location is a rich and growing tradition of cognitive neuroscience models and enquiry on working memory development. In the sections that follow, nosotros focus specifically on developmental cerebral neuroscience research on working retentiveness in infancy (for more than exhaustive reviews on memory development, meet Cowan, 1995; Nelson, 1995; Pelphrey and Reznick, 2003; Backbone and Howe, 2004; Rose et al., 2004; Bauer, 2009; Rovee-Collier and Cuevas, 2009).
Much of the research on working retention in infancy has focused on tasks similar to the Piagetian A-non-B task, and generally all tasks involve some delayed response (DR) with the correct response requiring some level of attentional control. The A-non-B and other DR tasks typically involve the presentation of 2 or more than wells. While the participant watches, an attractive object is placed in one of the wells and the participant'south view of the object is then occluded. Following a cursory delay, the participant is allowed to retrieve the object from i of the wells. In the A-not-B job, after multiple successful retrieval trials, the location of the subconscious object is reversed (again while the participant observes). The classic A-non-B fault occurs when the participant continues to reach for the object in the original hiding location after observing the reversal of the hiding location.
Diamond (1985, 1990) has attributed perseverative reaching on the A-not-B task to a lack of inhibitory control in younger participants and attributes higher success rates in older infants (8–9 months) to further maturation of dorsolateral prefrontal cortex (DLPFC). It has been noted (Diamond, 1990; Hofstadter and Reznick, 1996; Stedron et al., 2005) that participants occasionally look to the right location after reversal merely proceed to accomplish to the incorrect (previously rewarded) location. Hofstadter and Reznick (1996) found that when gaze and reach differ in direction, infants are more likely to direct their gaze to the correct location. Thus, poor operation in the A-not-B reaching chore may be influenced by immature inhibitory control of reaching behavior as opposed to a working retention deficiency. Alternatively, Smith et al. (1999) conducted a systematic series of experiments using the A-non-B task and establish that several factors other than inhibition contribute to perseverative reaching; including infant posture, direction of gaze, preceding activity, and long-term experiences in similar tasks. However, using an oculomotor version of the DR task, Gilmore and Johnson (1995) found that infants as young as 6 months of historic period were able to demonstrate successful performance. Similarly, using a peek-a-boo looking version of the DR chore, Reznick et al. (2004) found evidence of a developmental transition at around 6 months of age associated with improved working memory operation.
In several studies utilizing looking versions of the DR job, pregnant evolution has been found to occur from 5 to 12 months of age. With increasing historic period, infants testify higher rates of correct responses, and infants tin can tolerate longer delays and even so demonstrate successful responses (Hofstadter and Reznick, 1996; Pelphrey et al., 2004; Cuevas and Bell, 2010). Bell and colleagues (east.g., Bell and Adams, 1999; Bell, 2001, 2002, 2012; Bell and Wolfe, 2007; Cuevas and Bell, 2011) have integrated EEG measures in looking versions of the A-not-B task in a systematic line of piece of work on the development of working memory. Bell and Play tricks (1994) plant developmental change in baseline frontal EEG power was associated with functioning improvement on the A-not-B task. Power changes from baseline to task in the 6–ix Hz EEG frequency band also correlate with successful performance for viii-month-old infants (Bong, 2002). Additionally, higher levels of frontal-parietal and frontal-occipital EEG coherence every bit well every bit decreased center rate from baseline to chore are all associated with better performance on the looking version of the A-not-B task (Bell, 2012).
Taken together, these findings provide back up for the role of a frontal-parietal network in working memory tasks in infancy which is consistent with findings from neuroimaging studies with older children and adults showing recruitment of DLPFC, ventrolateral prefrontal cortex (VLPFC), intraparietal cortex, and posterior parietal cortex (Sweeney et al., 1996; Fuster, 1997; Courtney et al., 1997; D'Esposito et al., 1999; Klingberg et al., 2002; Crone et al., 2006; Scherf et al., 2006). For example, Crone et al. (2006) utilized fMRI during an object working retentivity task with children and adults and found that VLPFC was involved in maintenance processes for children and adults, and DLPFC was involved in manipulation of items in working memory for adults and children older than 12. The youngest group of children tested (8–12 years of historic period) did non recruit DLPFC during detail manipulation, and did not perform likewise as adolescents and adults on the task.
The change-detection chore is used to examine capacity limits for number of items an private can maintain in VSTM, and the coordinating alter-preference chore is used to measure capacity limits with infant participants. Similar to the VPC task, the change-preference job capitalizes on infants' trend to prefer novel or familiar stimuli. Two sets of stimuli are briefly and repeatedly presented to the left and right of midline with items in one fix of stimuli changing across each presentation and items in the other fix remaining constant. Infant looking to the left and correct stimulus set is measured and greater looking to the changing set up side is utilized as an index of working memory. Set size is manipulated to determine chapters limits for participants of different ages. Ross-Sheehy et al. (2003) institute a capacity increase from ane to iii items across vi.v–12.five months of age. The authors proposed that the increase in chapters limits on this task across this age range is driven in part past development of the ability to bind color to location. In a subsequent study, the authors (Ross-Sheehy et al., 2011) constitute that providing infants with an attentional cue facilitated memory for items in a stimulus set. Ten month-olds demonstrated enhanced functioning when provided with a spatial cue and v-calendar month-olds demonstrated enhanced performance when provided with a motility cue. These findings demonstrate that spatial orienting and selective attending influences infant performance on a VSTM chore, and back up the possibility that further development of the posterior orienting system influences maintenance processes involved in working memory in infancy.
Spencer and colleagues (e.g., Spencer et al., 2007; Simmering and Spencer, 2008; Simmering et al., 2008; Perone et al., 2011; Simmering, 2012) accept utilized dynamic neural field (DNF) models to explain developmental changes in the modify-preference chore. Using the DNF model, Perone et al. (2011) did simulation tests of the spatial precision hypothesis (SPH), predicting that the increased working memory capacity limits plant to develop during infancy are based on the strengthening of excitatory and inhibitory projections between a working memory field, perceptual field, and an inhibitory layer. According to the DNF model, the perceptual field consists of a population of neurons with receptive fields for certain characteristic dimensions (e.1000., color, shape), and activation in the working memory layer leads to inhibition of similarly tuned neurons in the perceptual field. The results of their simulation experiments were very similar to past behavioral findings and provided support for the SPH in explaining the increases in capacity limits that have been plant to occur with increasing age in infancy.
Findings from studies utilizing the change-preference task provide insight into capacity limits in VSTM during infancy. However, this task simply requires identification of novel items or objects based on maintenance of a retentiveness representation over very cursory delays (i.e., less than 500 ms). Given that delays between familiarization and testing on infant recognition memory tasks are typically very brief and the length of the filibuster is often not specified, information technology is particularly hard to determine whether or not recognition retentivity functioning is based on short-term memory or long-term memory. Call back that 4-month-olds only demonstrate recognition with upward to 10 s delays (Diamond, 1990). Thus, it is also difficult to make up one's mind whether or not operation on the change-preference task taps into maintenance of items in working retentivity or simply measures recognition memory. Alternatively, i could argue that performance on recognition retention tasks with brief delays may be driven past working memory. Interestingly, Perone and Spencer (2013a,b) again utilized the DNF model to simulate infant functioning on recognition memory tasks. The results of the simulations indicated that increasing the efficiency of excitatory and inhibitory interactions betwixt the perceptual field and a working retentivity field in their model led to novelty preferences on VPC trials with less exposure to the familiar stimulus. These faux results are similar to the developmental trends found to occur with increasing age across infancy in empirical studies utilizing the VPC task (due east.yard., Rose et al., 1982; Hunter and Ames, 1988; Freeseman et al., 1993). The authors concluded that development of working retentiveness is a pregnant factor in the increased likelihood that older infants volition demonstrate novelty preferences on recognition retentiveness tasks when compared to younger infants.
In guild to investigate working memory in infancy, Káldy and Leslie (2003, 2005) conducted a series of experiments with infants that involved both identification and individuation for successful operation. Individuation involves item or object identification combined with inbound the identified information into existing memory representations. Infants were familiarized with 2 objects of dissimilar shapes presented repeatedly in the middle of a phase. The side position of the objects was alternated across presentations in order to crave infants to integrate object shape with location on a trial past trial basis. During the examination phase, the objects were presented in the center of the stage as in familiarization and so placed behind occluders on the aforementioned side of the stage. After a delay, the occluders were removed. On change trials, removal of the occluders revealed that the unlike shaped objects were reversed in location. On no-change control trials, the objects remained in the same location upon removal of the occluders. Longer looking on change trials indicated individuation of the object based on identifying the alter in object shape from the location it was in prior to occlusion. Results indicated that while ix-month-olds could identify changes in object location for both objects (Káldy and Leslie, 2003), vi-month-olds were simply able to bind object to location for the final object that was moved behind the occluder in the examination phase (Káldy and Leslie, 2005). The authors ended that the younger infants' memory maintenance was more susceptible to distraction of attention. Káldy and Leslie (2005) also proposed that the significant improvements on this task between 6–ix months of age are related to further development of medial temporal lobe structures (i.e., enthorhinal cortex, parahippocampal cortex) which allows older infants to proceed to agree objects in working memory in the presence of distractors.
Thus, Káldy and Leslie (2003, 2005) and Káldy and Sigala (2004) have proposed an alternative model of working retentivity development which emphasizes the importance of medial temporal lobe structures more so than PFC. They argue that the majority of working memory models emphasizing the importance of DLPFC for working memory are confounding the response inhibition required in typical working memory tasks (due east.yard., the A-non-B task) with truthful working retentivity processes. To farther accost this limitation, Kaldy and colleagues (Káldy et al., 2015) designed a delayed match retrieval task which involves location-object binding but requires less response inhibition than the classic version of the A-not-B task. Infants are shown two cards, each with pictures of unlike objects or patterns on them. The cards are turned over and then a tertiary card is placed confront upwards which matches 1 of the face down cards. Infants are rewarded with an bonny stimulus for looks toward the location of the matching face down card. The authors tested 8- and 10-calendar month-olds on this task and found the 10-month-olds performed significantly higher up hazard levels. 8 month-olds performed at run a risk levels but showed improvement beyond trials. Thus, similar to previous work, significant gains in working memory performance are institute to occur in the second one-half of the kickoff postnatal year on the delayed friction match retrieval job.
Regarding Káldy and Sigala (2004) view that too much emphasis has been placed on the importance of PFC for infant working memory, results from the DNF simulations washed by Perone et al. (2011) also support the possibility that areas involved in visual processing and object recognition could account for successful working memory performance on the alter-preference task without requiring pregnant PFC contributions to attentional-control. However, in contempo exploratory studies utilizing functional nearly infrared spectroscopy (fNIRS) to measure out the BOLD response of infant participants during an object-permanence task. Baird et al. (2002) observed activation of frontal areas for babe participants during the chore. However, receptors were just applied to frontal sites, thus limiting the conclusion that the increased frontal action during this task was unique or of particular functional significance in comparison to other brain regions. Nonetheless, Buss et al. (2014) utilized fNIRS to image cortical activeness associated with visual working memory capacity in iii- and 4-year-old children. In this study, receptors were applied over frontal and parietal locations. Frontal and parietal channels in the left hemisphere showed increased activation when working memory load was increased from one to 3 items. Results supported the possibility that immature children use a frontal-parietal working retentivity excursion similar to adults. Both of these findings from fNIRS studies provide preliminary back up for the role of PFC in working retentiveness during early evolution.
Luciana and Nelson (1998) emphasize the critical office the PFC plays in integrating sensorimotor traces in working memory to guide future behavior. According to Luciana and Nelson, the A-not-B task may actually overestimate the functional maturity of the PFC in babe participants considering it does not require the accurate integration of sensorimotor traces in working retentiveness. They propose the integration of sensorimotor traces should be considered a core procedure in working memory definitions. The majority of working memory definitions include executive control components, and persistent activity in DLPFC has been linked with control functions involved in the manipulation of information for the purpose of goal-directed activity (eastward.one thousand., Curtis and D'Esposito, 2003; Crone et al., 2006). Thus, the exact contribution of PFC to working memory functions in early on development remains unclear. What is clear from the extant literature is that infants beyond 5–6 months of age are capable of demonstrating bones yet immature aspects of working memory, and meaning improvement in these basic functions occurs from v–half dozen months (e.g., Diamond, 1990; Gilmore and Johnson, 1995; Hofstadter and Reznick, 1996; Káldy and Leslie, 2003, 2005; Káldy and Sigala, 2004; Pelphrey et al., 2004; Reznick et al., 2004; Cuevas and Bell, 2010).
The Evolution of Attention Systems and Working Retentivity
Like to recognition memory, the improvements in working memory performance which occur subsequently 5–six months of age are likely influenced by further development of the attention systems previously discussed. The majority of the working retention studies discussed above examined visuospatial working memory. Operation on all of these working memory tasks involves voluntary eye movements and controlled scanning of the stimuli involved in the task. Thus, functional maturity of the posterior orienting organisation would be primal for successful performance on these tasks. This organisation shows pregnant development from 3 to 6 months of historic period (Johnson et al., 1991; Colombo, 2001; Courage et al., 2006; Reynolds et al., 2013). This timing coincides with the fourth dimension frame at which infants begin to demonstrate higher up take chances operation on working retentiveness tasks. For case, Gilmore and Johnson (1995) reported successful performance on an oculomotor DR task for half dozen-month-former infants, and Reznick et al. (2004) draw six months of age as a fourth dimension of transition for operation on a peek-a-boo version of the DR chore.
Successful functioning on working retentivity tasks involves more than than just voluntary control of eye movements. Working memory tasks also involve attentional control and inhibition. These cognitive functions are both associated with the anterior attending system (Posner and Peterson, 1990), which shows meaning and protracted development from 6 months on. Several studies have shown significant improvement on DR and change-preference tasks from 5 to 12 months of age (Hofstadter and Reznick, 1996; Ross-Sheehy et al., 2003; Pelphrey et al., 2004; Cuevas and Bell, 2010), an age range that overlaps with the functional onset of the anterior attention system. Given that some models emphasize the office of PFC and attentional control as existence disquisitional for working memory (eastward.g., Baddeley, 1996; Kane and Engle, 2002; Klingberg et al., 2002), further development of the anterior attention system would be disquisitional for working memory development (for further discussion of attention and memory relations in childhood and adulthood, see Awh and Jonides, 2001; Awh et al., 2006; Astle and Scerif, 2011; Amso and Scerif, 2015).
The general arousal/attending organisation shows significant developmental change across infancy and early on childhood characterized by gains in both the magnitude and duration of periods of sustained attending (Richards and Cronise, 2000; Richards and Turner, 2001; Reynolds and Richards, 2008). Infants are more than likely to demonstrate testify of recognition retentiveness if initial exposure to the test stimulus occurs during sustained attending or if the infant is engaged in sustained attending during the recognition test (eastward.g., Richards, 1997; Frick and Richards, 2001; Reynolds and Richards, 2005; Reynolds et al., 2010). Information technology stands to reason that these developmental gains in sustained attention would also facilitate improved performance on working memory tasks. This reasoning is supported by Bong (2012) finding that infants who show decreased heart rate from baseline to task too show enhanced performance on the A-non-B task. Studies utilizing the heart rate phases (Richards and Casey, 1992) during infant working memory tasks would provide greater insight into the effects of sustained attending on working memory operation.
Relations between arousal and attending are complex and change throughout development. The pregnant and sustained decrease in heart charge per unit associated with attention is near likely limited to infancy and early on childhood; all the same, individual differences in center rate variability are related to attention and cognitive functioning throughout development (Porges, 1992; Suess et al., 1994; Reynolds and Richards, 2008). Relatively little work has examined the influence of arousal aspects of attending on working retention in later development. An exception would be the work past Thayer and colleagues (Hansen et al., 2003; Thayer et al., 2009) examining relations between HRV and working memory in adults. Their findings bespeak that individual differences in baseline HRV are associated with operation on working memory tasks. Individuals with high baseline HRV perform amend on working memory tasks than individuals with depression baseline HRV, and the advantage is specific to tasks requiring executive function (Thayer et al., 2009). Thus, attention and arousal appear to influence working memory throughout development; however, the dynamics of these relations are circuitous and would be expected to change significantly with age.
The development of attention and the development of working memory are closely related. Meaning gains on working memory tasks overlap in developmental timing with primal periods for development of sustained attending, the posterior orienting organization, and the anterior attention system. There is also significant overlap in neural systems involved in attention and working memory. The cortical sources of the Nc ERP component associated with infant visual attention accept been localized to areas of PFC (Reynolds and Richards, 2005; Reynolds et al., 2010). Similarly, research with fNIRS indicates that frontal and parietal areas are involved in working retentivity performance for infants (Baird et al., 2002) and preschoolers (Buss et al., 2014). Given the substantial overlap in developmental timing and neural systems involved in both attention and working memory, future research should aim to examine relations between attention and working memory in infancy and early babyhood using both psychophysiological and neural measures. A multi-level analysis approach would exist platonic for addressing the controversy regarding the relative contribution of PFC, parietal cortex, and medial temporal lobe structures to working memory operation. Attending plays a key part in successful working memory performance, and the development of attending systems most likely influences the development of working memory. Bidirectional effects are common throughout development, and thus of equal interest is the potential influence of working memory on farther development of attention systems in infancy and early babyhood.
Author Contributions
After discussions well-nigh potential directions for the commodity, the authors (Gdr and ACR) settled on the overall content to include and outline to follow for the article. ACR provided recommendations on potential content for several of the major sections of the commodity. GDR incorporated much of ACR's work into the commodity when he wrote the original draft, and subsequently incorporated farther input from ACR into the terminal version of the manuscript.
Conflict of Involvement Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could exist construed as a potential disharmonize of involvement.
Acknowledgments
Inquiry reported in this commodity and the writing of this article were supported by the National Constitute of Kid Health and Human Development Grant R21-HD065042, and the National Science Foundation Developmental and Learning Sciences Division Grant 1226646 to GDR.
References
Astle, D. Due east., and Scerif, M. (2011). Interactions betwixt attention and visual short-term memory (VSTM): what can exist learnt from private and developmental differences? Neuropsychologia 49, 1435–1445. doi: x.1016/j.neuropsychologia.2010.12.001
PubMed Abstruse | CrossRef Full Text | Google Scholar
Bachevalier, J., Brickson, Thou., and Hagger, C. (1993). Limbic-dependent recognition memory in monkeys develops early in infancy. Neuroreport 4, 77–80. doi: 10.1097/00001756-199301000-00020
PubMed Abstract | CrossRef Total Text | Google Scholar
Baird, A. A., Kagan, J., Gaudette, T., Walz, K. A., Hershlag, N., and Boas, D. A. (2002). Frontal lobe activation during object permanence: data from near-infrared spectroscopy. Neuroimage 16, 1120–1126. doi: 10.1006/nimg.2002.1170
CrossRef Full Text | Google Scholar
Bauer, P. J. (2009). "The cognitive neuroscience of the development of memory," in The Development of Retention in Infancy and Babyhood, eds M. Courage and N. Cowan (New York, NY: Psychology Press), 115–144.
Google Scholar
Begleiter, H., Porjesz, B., and Wang, West. (1993). A neurophysiologic correlate of visual short-term retentivity in humans. Electroencephalogr. Clin. Neurophysiol. 87, 46–53. doi: 10.1016/0013-4694(93)90173-s
PubMed Abstract | CrossRef Full Text | Google Scholar
Bell, Chiliad. A. (2001). Brain electric activity associated with cognitive processing during a looking version of the A-non-B chore. Infancy 2, 311–330. doi: 10.1207/s15327078in0203_2
CrossRef Full Text | Google Scholar
Bell, M. A., and Adams, S. E. (1999). Comparable performance on looking and reaching versions of the A-non-B task at 8 months of age. Infant Behav. Dev. 22, 221–235. doi: 10.1016/s0163-6383(99)00010-7
CrossRef Full Text | Google Scholar
Bell, M. A., and Trick, N. A. (1994). "Brain development over the first twelvemonth of life: relations between EEG frequency and coherence and cognitive and melancholia behaviors," in Human Behavior and the Developing Brain, eds G. Dawson and K. Fischer (New York, NY: Guilford), 314–345.
Google Scholar
Bell, M. A., and Wolfe, C. D. (2007). Changes in brain functioning from infancy to early childhood: show from EEG power and coherence during working memory tasks. Dev. Neuropsychol. 31, 21–38. doi: 10.1207/s15326942dn3101_2
PubMed Abstract | CrossRef Full Text | Google Scholar
Buss, A. T., Fox, N., Boas, D. A., and Spencer, J. P. (2014). Probing the early on evolution of visual working memory capacity with functional near-infrared spectroscopy. Neuroimage 85, 314–325. doi: 10.1016/j.neuroimage.2013.05.034
PubMed Abstract | CrossRef Full Text | Google Scholar
Colombo, J., and Mitchell, D. W. (1990). "Individual and developmental differences in infant visual attending," in Private Differences in Infancy, eds J. Colombo and J. W. Fagen (Hillsdale, NJ: Erlbaum), 193–227.
Courage, M. L., and Howe, M. L. (2004). Advances in early memory development research: insights about the dark side of the moon. Dev. Rev. 24, half-dozen–32. doi: ten.1016/j.dr.2003.09.005
CrossRef Total Text | Google Scholar
Courage, M. L., Reynolds, M. D., and Richards, J. East. (2006). Infants' attention to patterned stimuli: developmental change from iii to 12 months of age. Child Dev. 77, 680–695. doi: 10.1111/j.1467-8624.2006.00897.x
PubMed Abstruse | CrossRef Total Text | Google Scholar
Courtney, Due south. M., Ungerleider, L. G., Keil, K., and Haxby, J. V. (1997). Transient and sustained activity in a distributed neural system for human working retention. Nature 386, 608–611. doi: ten.1038/386608a0
PubMed Abstract | CrossRef Full Text | Google Scholar
Cowan, Northward. (1995). Attention and Retention: An Integrated Framework. New York, NY: Oxford University Press.
Crone, E. A., Wendelken, C., Donohue, Southward., van Leijenhorst, L., and Bunge, S. A. (2006). Neurocognitive development of the power to dispense information in working retentivity. Proc. Natl. Acad. Sci. U South A 103, 9315–9320. doi: ten.1073/pnas.0510088103
PubMed Abstract | CrossRef Total Text | Google Scholar
Cuevas, K., and Bell, M. A. (2011). EEG and ECG from 5 to x months of age: developmental changes in baseline activation and cognitive processing during a working memory task. Int. J. Psychophysiol. eighty, 119–128. doi: 10.1016/j.ijpsycho.2011.02.009
PubMed Abstract | CrossRef Full Text | Google Scholar
de Haan, M., and Nelson, C. A. (1997). Recognition of the female parent's face by six-month-onetime infants: a neurobehavioral written report. Kid Dev. 68, 187–210. doi: 10.1111/j.1467-8624.1997.tb01935.ten
PubMed Abstract | CrossRef Full Text | Google Scholar
de Haan, M., and Nelson, C. A. (1999). Encephalon activity differentiates face and object processing in half dozen-calendar month-old infants. Dev. Psychol. 35, 1113–1121. doi: 10.1037/0012-1649.35.iv.1113
PubMed Abstract | CrossRef Full Text | Google Scholar
D'Esposito, M., Postle, B. R., Ballard, D., and Lease, J. (1999). Maintenance versus manipulation of information held in working retentivity: an event-related fMRI study. Brain cogn. 41, 66–86. doi: 10.1006/brcg.1999.1096
CrossRef Total Text | Google Scholar
Diamond, A. (1990). "Rate of maturation of the hippocampus and the developmental progression of children'southward performance on the delayed non-matching to sample and visual paired comparison tasks," in Development and Neural Bases of Higher Cognitive Functions, ed. A. Diamond (New York, NY: New York Academy of Sciences Press), 394–426.
Google Scholar
Eichenbaum, H., Yonelinas, A., and Ranganath, C. (2007). The medial temporal lobe and recognition retentivity. Annu. Rev. Neurosci. xxx, 123–152. doi: 10.1146/annurev.neuro.30.051606.094328
PubMed Abstract | CrossRef Total Text | Google Scholar
Fahy, F. L., Riches, I. P., and Brownish, M. Westward. (1993). Neuronal activity related to visual recognition memory: long-term memory and the encoding of recency and familiarity information in the primate anterior and medial inferior rhinal cortex. Exp. Encephalon Res. 96, 457–472. doi: 10.1007/bf00234113
PubMed Abstruse | CrossRef Full Text | Google Scholar
Freeseman, L. J., Colombo, J., and Coldren, J. T. (1993). Individual differences in infant visual attending: iv-month-olds' discrimination and generalization of global and local stimulus properties. Kid Dev. 64, 1191–1203. doi: 10.2307/1131334
PubMed Abstract | CrossRef Full Text | Google Scholar
Frick, J. E., and Richards, J. East. (2001). Individual differences in infants' recognition of briefly presented visual stimuli. Infancy 2, 331–352. doi: 10.1207/s15327078in0203_3
CrossRef Full Text | Google Scholar
Fuster, J. M. (1997). The Prefrontal Cortex: Beefcake, Physiology, and Neuropsychology of the Frontal Lobes. New York: Raven Press.
Google Scholar
Gilmore, R., and Johnson, 1000. H. (1995). Working retentiveness in infancy: 6-month-olds′ performance on two versions of the oculomotor delayed response job. J. Exp. Child Psychol. 59, 397–418. doi: x.1006/jecp.1995.1019
PubMed Abstract | CrossRef Full Text | Google Scholar
Guy, M. W., Reynolds, Yard. D., and Zhang, D. (2013). Visual attending to global and local stimulus backdrop in 6-month-old infants: private differences and event-related potentials. Child Dev. 84, 1392–1406. doi: x.1111/cdev.12053
PubMed Abstract | CrossRef Total Text | Google Scholar
Guy, M. W., Zieber, Northward., and Richards, J. E. (in press). The cortical evolution of specialized face processing in infancy. Child Dev. 84, 1392–1406.
Hood, B. M. (1995). Shifts of visual attention in the human infant: a neuroscientific arroyo. Adv. Infancy Res. x, 163–216.
Hunter, One thousand., and Ames, E. (1988). "A multifactor model of infant preferences for novel and familiar stimuli," in Advances in Infancy Research, (Vol. five), eds C. Rovee-Collier and Fifty. P. Lipsitt (Norwood, NJ: Ablex), 69–95.
Google Scholar
Johnson, K. H., Posner, M., and Rothbart, M. One thousand. (1991). Components of visual orienting in early infancy: contingency learning, anticipatory looking and disengaging. J. Cogn. Neurosci. 3, 335–344. doi: 10.1162/jocn.1991.3.4.335
PubMed Abstruse | CrossRef Full Text | Google Scholar
Káldy, Z., Guillory, South., and Blaser, E. (2015). Delayed friction match retrieval: a novel anticipation-based visual working memory prototype. Dev. Sci. doi: 10.1111/desc.12335 [Epub ahead of print]
PubMed Abstract | CrossRef Full Text | Google Scholar
Káldy, Z., and Leslie, A. M. (2003). Identification of objects in 9-month-old infants: integrating 'what'and 'where'information. Dev. Sci. 6, 360–373. doi: 10.1111/1467-7687.00290
CrossRef Full Text | Google Scholar
Káldy, Z., and Sigala, N. (2004). The neural mechanisms of object working memory: what is where in the infant brain? Neurosci. Biobehav. Rev. 28, 113–121. doi: 10.1016/j.neubiorev.2004.01.002
PubMed Abstruse | CrossRef Full Text | Google Scholar
Kane, M. J., and Engle, R. W. (2002). The role of prefrontal cortex in working-memory capacity, executive attending and general fluid intelligence: an individual differences perspective. Psychon. Bull. Rev. ix, 637–671. doi: 10.3758/bf03196323
PubMed Abstract | CrossRef Full Text | Google Scholar
Klingberg, T., Forssberg, H., and Westerberg, H. (2002). Increased encephalon activity in frontal and parietal cortex underlies the evolution of visuospatial working memory capacity in babyhood. J. Cogn. Neurosci. 14, ane–10. doi: x.1162/089892902317205276
PubMed Abstract | CrossRef Total Text | Google Scholar
Li, Fifty., Miller, E. K., and Desimone, R. (1993). The representation of stimulus familiarity in anterior inferior temporal cortex. J. Neurophysiol. 69, 1918–1929.
PubMed Abstruse | Google Scholar
Luciana, M., and Nelson, C. A. (1998). The functional emergence of prefrontally-guided memory systems in four- to eight-twelvemonth-former children. Neuropsychologia 36, 272–293. doi: 10.1016/s0028-3932(97)00109-7
PubMed Abstruse | CrossRef Full Text | Google Scholar
Nelson, C. A. (1995). The ontogeny of human being retention: a cognitive neuroscience perspective. Developmental Psychology 5, 723–738. doi: x.1002/9780470753507.ch10
CrossRef Total Text | Google Scholar
Pelphrey, Grand. A., and Reznick, J. Due south. (2003). "Working memory in infancy," in Advances in Child Beliefs, (Vol. 31), ed. R. V. Kail (San Diego, CA: Academic Press), 173–227.
Google Scholar
Pelphrey, K. A., Reznick, J. S., Davis Goldman, B., Sasson, North., Morrow, J., Donahoe, A., et al. (2004). Evolution of visuospatial brusk-term memory in the second half of the 1st year. Dev. Psychol. 40, 836–851. doi: 10.1037/0012-1649.forty.5.836
PubMed Abstract | CrossRef Full Text | Google Scholar
Perone, S., Simmering, V., and Spencer, J. (2011). Stronger neural dynamics capture changes in infants' visual working memory capacity over development. Dev. Sci. xiv, 1379–1392. doi: 10.1111/j.1467-7687.2011.01083.ten
PubMed Abstruse | CrossRef Full Text | Google Scholar
Perone, Southward., and Spencer, J. P. (2013a). Autonomy in action: linking the deed of looking to retention formation in infancy via dynamic neural fields. Cogn. Sci. 37, 1–60. doi: x.1111/cogs.12010
PubMed Abstract | CrossRef Total Text | Google Scholar
Perone, Due south., and Spencer, J. P. (2013b). Democratic visual exploration creates developmental modify in familiarity and novelty seeking behaviors. Front. Psychol. four:648. doi: 10.3389/fpsyg.2013.00648
PubMed Abstract | CrossRef Full Text | Google Scholar
Porges, S. W. (1992). "Autonomic regulation and attending," in Attention and Information Processing in Infants and Adults: Perspectives from Human and Creature Research, eds B. A. Campbell, H. Hayne, and R. Richardson (Hillsdale, NJ: Lawrence Erlbaum Assembly), 201–223.
Google Scholar
Reynolds, G. D., Courage, 1000. 50., and Richards, J. East. (2010). Babe attention and visual preferences: converging evidence from behavior, event-related potentials and cortical source localization. Dev. Psychol. 46, 886–904. doi: 10.1037/a0019670
PubMed Abstruse | CrossRef Total Text | Google Scholar
Reynolds, G. D., Backbone, M. L., and Richards, J. E. (2013). "The development of attention," in Oxford Handbook of Cerebral Psychology, ed. D. Reisberg (New York, NY: Oxford University Press), 1000–1013.
Google Scholar
Reynolds, G. D., Guy, K. Due west., and Zhang, D. (2011). Neural correlates of individual differences in baby visual attention and recognition memory. Infancy sixteen, 368–391. doi: x.1111/j.1532-7078.2010.00060.x
CrossRef Total Text | Google Scholar
Reynolds, G. D., and Richards, J. E. (2005). Familiarization, attention and recognition memory in infancy: an ERP and cortical source localization study. Dev. Psychol. 41, 598–615. doi: 10.1037/0012-1649.41.iv.598
PubMed Abstract | CrossRef Total Text | Google Scholar
Reynolds, Yard. D., and Richards, J. E. (2008). "Infant center rate: a developmental psychophysiological perspective," in Developmental Psychophysiology: Theory, Systems and Applications, eds L. A. Schmidt and Due south. J. Segalowitz (Cambridge: Cambridge University Printing), 173–212.
Reznick, J. S., Morrow, J. D., Goldman, B. D., and Snyder, J. (2004). The onset of working memory in infants. Infancy half-dozen, 145–154. doi: 10.1207/s15327078in0601_7
CrossRef Full Text | Google Scholar
Richards, J. E. (1985). The development of sustained visual attention in infants from xiv to 26 weeks of age. Psychophysiology 22, 409–416. doi: 10.1111/j.1469-8986.1985.tb01625.x
CrossRef Total Text | Google Scholar
Richards, J. E. (1997). Effects of attention on infants' preference for briefly exposed visual stimuli in the paired-comparison recognition-memory paradigm. Dev. Psychol. 33, 22–31. doi: 10.1037/0012-1649.33.1.22
PubMed Abstract | CrossRef Full Text | Google Scholar
Richards, J. East. (2008). "Attending in young infants: a developmental psychophysiological perspective," in Handbook of Developmental Cognitive Neuroscience, eds C. A. Nelson and M. Luciana (Cambridge, MA: MIT Press), 479–497.
Google Scholar
Richards, J. East. (2010). "Attending in the brain and early on infancy," in Neoconstructivism: The New Science of Cognitive Development, ed. Due south. P. Johnson (New York, NY: Oxford Academy Press), 3–31.
Google Scholar
Richards, J. E., and Casey, B. J. (1992). "Development of sustained visual attention in the human infant," in Attention and Data Processing in Infants and Adults: Perspectives from Man and Creature Inquiry, eds B. A. Campbell and H. Hayne (Hillsdale, NJ: Erlbaum Publishing), 30–threescore.
Google Scholar
Richards, J. E., and Cronise, K. (2000). Extended visual fixation in the early preschool years: look duration, center rate changes and attentional inertia. Child Dev. 71, 602–620. doi: x.1111/1467-8624.00170
PubMed Abstract | CrossRef Total Text | Google Scholar
Richards, J. East., and Turner, Eastward. D. (2001). Extended visual fixation and distractibility in children from six to twenty-four months of historic period. Child Dev. 72, 963–972. doi: 10.1111/1467-8624.00328
PubMed Abstract | CrossRef Full Text | Google Scholar
Rose, S. A., Feldman, J. F., and Jankowski, J. J. (2004). Infant visual recognition retentivity. Dev. Rev. 24, 74–100. doi: 10.1016/j.doc2003.09.004
CrossRef Full Text | Google Scholar
Rose, S. A., Gottfried, A. W., Melloy-Carminar, P. Thousand., and Bridger, W. H. (1982). Familiarity and novelty preferences in infant recognition retentivity: implications for information processing. Dev. Psychol. 18, 704–713. doi: 10.1037/0012-1649.18.v.704
CrossRef Full Text | Google Scholar
Ross-Sheehy, Due south., Oakes, L. Thousand., and Luck, Southward. J. (2003). The development of visual short-term retention capacity in infants. Child Dev. 74, 1807–1822. doi: 10.1046/j.1467-8624.2003.00639.x
PubMed Abstruse | CrossRef Total Text | Google Scholar
Ross-Sheehy, S., Oakes, Fifty. M., and Luck, Due south. J. (2011). Exogenous attention influences visual brusk-term memory in infants. Dev. Sci. 14, 490–501. doi: 10.1111/j.1467-7687.2010.00992.10
PubMed Abstract | CrossRef Full Text
Rovee-Collier, C., and Cuevas, K. (2009). Multiple memory systems are unnecessary to account for baby memory evolution: an ecological model. Dev. Psychol. 45, 160-174. doi: 10.1037/a0014538
PubMed Abstruse | CrossRef Full Text | Google Scholar
Ruff, H. A., and Rothbart, Grand. Yard. (1996). Attending in Early Development. New York: Oxford University Press.
Google Scholar
Sarter, Yard., Givens, B., and Bruno, J. P. (2001). The cognitive neuroscience of sustained attention: where pinnacle-downwards meets lesser-upwards. Brain Res. Encephalon Res. Rev. 35, 146–160. doi: 10.1016/s0165-0173(01)00044-3
PubMed Abstruse | CrossRef Total Text | Google Scholar
Scherf, Grand. S., Sweeney, J. A., and Luna, B. (2006). Brain basis of developmental modify in visuospatial working memory. J. Cogn. Neurosci. 18, 1045–1058. doi: ten.1162/jocn.2006.18.7.1045
PubMed Abstract | CrossRef Full Text | Google Scholar
Schiller, P. H. (1985). "A model for the generation of visually guided saccadic middle movements," in Models of the Visual Cortex, eds D. Rose and V. G. Dobson (New York, NY: Wiley), 62–seventy.
Google Scholar
Simmering, V. R. (2012). The development of visual working memory capacity during early on childhood. J. Exp. Child. Psychol. 111, 695–707. doi: ten.1016/j.jecp.2011.10.007
CrossRef Total Text | Google Scholar
Simmering, V. R., Schutte, A. R., and Spencer, J. P. (2008). Generalizing the dynamic field theory of spatial noesis across real and developmental time scales. Brain Res. 1202, 68–86. doi: 10.1016/j.brainres.2007.06.081
CrossRef Full Text | Google Scholar
Simmering, V. R., and Spencer, J. P. (2008). Generality with specificity: the dynamic field theory generalizes across tasks and time scales. Dev. Sci. xi, 541–555. doi: 10.1111/j.1467-7687.2008.00700.x
CrossRef Full Text | Google Scholar
Smith, L. B., Thelen, Due east., Titzer, R., and McLin, D. (1999). Knowing in the context of acting: the job dynamics of the A-not-B error. Psychol. Rev. 106, 235-260. doi: 10.1037/0033-295x.106.2.235
PubMed Abstract | CrossRef Full Text | Google Scholar
Snyder, K. (2010). Neural correlates of encoding predict infants' retentiveness in the paired-comparison process. Infancy xv, 270–299. doi: x.1111/j.1532-7078.2009.00015.x
CrossRef Full Text | Google Scholar
Sokolov, East. N. (1963). Perception and the Conditioned Reflex. Oxford: Pergamon Press.
Google Scholar
Spencer, J. P., Simmering, V. R., Schutte, A. R., and Schöner, Grand. (2007). "What does theoretical neuroscience have to offering the study of behavioral evolution? Insights from a dynamic field theory of spatial cognition," in The Emerging Spatial Mind, eds J. Plumert and J. P. Spencer (New York, NY: Oxford Academy Press), 320–321.
Stedron, J. Thou., Sahni, S. D., and Munakata, Y. (2005). Mutual mechanisms for working memory and attending: the case of perseveration with visible solutions. J. Cogn. Neurosci. 17, 623–631. doi: 10.1162/0898929053467622
PubMed Abstract | CrossRef Full Text | Google Scholar
Suess, P. East., Porges, S. W., and Plude, D. J. (1994). Cardiac vagal tone and sustained attention in school-age children. Psychophysiology 31, 17–22. doi: 10.1111/j.1469-8986.1994.tb01020.ten
CrossRef Full Text | Google Scholar
Sweeney, J. A., Mintun, M. A., Kwee, S., Wiseman, 1000. B., Brown, D. L., Rosenberg, D. R., et al. (1996). Positron emission tomography written report of voluntary saccadic heart movements and spatial working memory. J. Neurophysiol. 75, 454–468.
PubMed Abstract | Google Scholar
Thayer, J. F., Hansen, A. L., Saus-Rose, E., and Johnsen, B. H. (2009). Heart charge per unit variability, prefrontal neural part and cognitive performance: the neurovisceral integration perspective on self-regulation, adaptation and health. Ann. Behav. Med. 37, 141–153. doi: 10.1007/s12160-009-9101-z
PubMed Abstruse | CrossRef Full Text | Google Scholar
Wan, H., Aggleton, J. P., and Brown, M. W. (1999). Different contributions of the hippocampus and perirhinal cortex to recognition retentivity. J. Neurosci. 19, 1142–1148.
PubMed Abstract | Google Scholar
Wiggs, C. L., and Martin, A. (1998). Properties and mechanisms of perceptual priming. Curr. Opin. Neurobiol. 8, 227–233. doi: 10.1016/S0959-4388(98)80144-10
CrossRef Full Text | Google Scholar
Xiang, J.-Z., and Dark-brown, M. W. (1998). Differential neuronal encoding of novelty, familiarity and recency in regions of the anterior temporal lobe. Neuropharmacology 37, 657–676. doi: x.1016/s0028-3908(98)00030-6
PubMed Abstract | CrossRef Total Text | Google Scholar
Zeamer, A., Heuer, Eastward., and Bachevalier, J. (2010). Developmental trajectory of object recognition in baby rhesus macaques with and without neonatal hippocampal lesions. J. Neurosci. 30, 9157–9165. doi: ten.1523/JNEUROSCI.0022-10.2010
PubMed Abstract | CrossRef Full Text | Google Scholar
Zhu, Ten. O., Brown, Thousand. Westward., McCabe, B. J., and Aggleton, J. P. (1995). Effects of the novelty or familiarity of visual stimuli on the expression of the intermediate early cistron c-fos in the rat brain. Neuroscience 69, 821–829. doi: 10.1016/0306-4522(95)00320-i
PubMed Abstract | CrossRef Full Text | Google Scholar
Source: https://www.frontiersin.org/articles/10.3389/fnsys.2016.00015/full
0 Response to "Memory Improves Dramatically Between 1 and 2 Months of Age and Then Again by 36 Months"
Post a Comment